O projekcie siatkówki kręgowców
Twierdzi się powszechnie, że oko kręgowców nie jest optymalne, jeśli chodzi o funkcjonowanie, ponieważ fotoreceptory w siatkówce są skierowane w kierunku przeciwnym niż padające światło. Jednak istnieją bardzo ważne funkcjonalne powody, by fotoreceptory kręgowców były tak właśnie skierowane. Struktura i funkcja fotoreceptorów zależy od ważnej warstwy, barwnego nabłonka siatkówki (RPE), który ponownie używa zużyte fotobarwniki, usuwa zużyte zewnętrzne części fotoreceptorów, dostarcza nieprzezroczystej warstewki absorbującej nadmierne światło i pełni dodatkowe funkcje. Te aspekty struktury i funkcjonowania oka kręgowców były ignorowane w argumentach ewolucjonistycznych na temat nieoptymalności, ale są istotne, by zrozumieć, jak oko funkcjonuje.
Wprowadzenie: popularny argument[1]
Argumentowano obszernie zarówno w technicznej,[2] jak i w popularnej literaturze ewolucjonistycznej,[3] że oko kręgowców jest kiepsko zaprojektowane. „Faktycznie jest ono zaprojektowane głupio” — napisał wpływowy teoretyk neodarwinizmu, George Williams — „ponieważ występuje w nim wiele funkcjonalnie arbitralnych i nieprzystosowujących cech”.[4] Według Williamsa głównie chodzi o odwrócenie siatkówki.[5]
„Siatkówka jest skierowana dolną stroną do góry” — pisze on. — „Pręciki i czopki stanowią dolną warstwę, w rezultacie czego światło dociera do nich dopiero po przejściu przez nerwy i naczynia krwionośne”. Według biologa ewolucyjnego z Uniwersytetu Kalifornijskiego w Los Angeles, Jareda Diamonda, struktury te
nie zostały umiejscowione za fotoreceptorami, gdzie każdy rozsądny inżynier by je umieścił, ale przed nimi, gdzie zasłaniają do pewnego stopnia padające światło. Projektant aparatu fotograficznego, który popełniłby taką gafę, zostałby bez zwłoki zwolniony z pracy.[6]
Argument ten kończy się stwierdzeniem, że siatkówka głowonogów (kałamarnic i ośmiornic) jest prawdopodobnie „poprawnie zainstalowana”, gdyż jej fotoreceptory skierowane są ku światłu, a nerwy „starannie schowane za warstwą fotoreceptorów”.[7] Mówi się, że głowonogi mają siatkówkę taką, jak trzeba.
Zastanawiając się nad tym argumentem możemy od razu przestać porównywać projekty siatkówki głowonogów (bezkręgowców) oraz kręgowców. Żaden z cytowanych autorów nie dostarczył świadectwa, że siatkówka głowonogów przewyższa co do funkcjonowania siatkówkę kręgowców: twierdzenie takie w każdym razie wydawałoby się nierozsądne. Czy setki tysięcy gatunków kręgowców, żyjących w bardzo różnych naziemnych, morskich i powietrznych środowiskach, naprawdę widziałoby lepiej, gdyby używały one układu wzrokowego używanego przez garstkę wyłącznie morskich bezkręgowców? Przy braku ścisłego materiału porównawczego wszelkie twierdzenia, że siatkówka głowonogów przewyższa funkcjonalnie siatkówkę kręgowców, pozostają wyłącznie hipotezami. Krótko mówiąc, nie ma powodu, by w nie wierzyć.
Ale należy na sprawę spojrzeć z bardziej zasadniczego punktu widzenia. Dlaczego w ogóle odnosimy się do siatkówki głowonogów? Jeśli ktoś twierdzi, że głowonogi mają właściwie zbudowaną siatkówkę, to zakłada, że kręgowce takiej nie mają i że robią, co mogą, w tej niekorzystnej sytuacji. Ale, oczywiście, należy jeszcze wykazać, że faktycznie siatkówka kręgowców jest niedoskonała. A tego nie wykazali ani cytowani wyżej autorzy, ani inni biologowie ewolucyjni. „Jedna z trudności, na jakie napotyka hipoteza optymalności” — zwracają uwagę Farnsworth i Niklas „polega na dostępie do obserwacji, które by ją przetestowały”.[8] Dotyczy to także hipotezy niedoskonałości, czego przykładem jest literatura ewolucjonistyczna na temat siatkówki kręgowców. Świat biologii jest pełen zagadkowych systemów. Niełatwo zrozumieć, dlaczego fotoreceptory kręgowców odwracają się tyłem do światła, ani dlaczego warstwy innych komórek znajdują się między fotoreceptorami i światłem. Ale bardzo wiele rzeczy w nauce nie jest oczywistych na pierwszy rzut oka. Trzeba przyjrzeć się dokładniej.
W tym przypadku nie musimy patrzeć daleko. Istnieją poważne funkcjonalne powody, by fotoreceptory kręgowców były tak skierowane, jak są. Te aspekty struktury i funkcjonowania siatkówki są ignorowane w ewolucjonistycznych argumentach na temat niedoskonałości, jednak są istotne dla zrozumienia, jak funkcjonuje oko.
Struktura siatkówki kręgowców
Najpierw trochę anatomii. Rysunek 1 przedstawia przekrój oka kręgowców. Światło przechodzi najpierw przez rogówkę, pierwszy element skupiający światło, potem przez źrenicę, która kontroluje, jak dużo światła będzie docierało do oka, i w końcu przez soczewkę, która pozwala regulować zogniskowanie światła. Światło o wyregulowanej intensywności zogniskowane jest na cienkiej warstwie, wyściełającej dno oka: siatkówce.
Siatkówka (patrz rys. 2) zbudowana jest z komórek centralnego układu nerwowego (CUN) i przekształca światło w impulsy elektryczne, właściwe dla CUN. Jest to bardzo skomplikowana warstwa, gdyż siatkówka zawiera komórki kilku różnych typów:
Fotoreceptory (pręciki i czopki, zaznaczone jako R i C na rys. 2), które faktycznie przekształcają energie światła w impuls elektryczny. Są to pierwsze komórki bezpośrednio zaangażowane w przenoszenie informacji w układzie wzrokowym, przesyłanie impulsów przez synapsę chemiczną do:
komórek dwubiegunowych (B na rysunku), następnej klasy komórek w siatkówce; komórki dwubiegunowe łączą się z:
komórkami zwojowymi (G na rysunku), wewnętrznej powierzchni siatkówki; komórki te mają aksony, które przemieszczają się razem, opuszczając oko przez tarczę nerwu wzrokowego i formując na drodze do mózgu nerw wzrokowy.
Komórki amakrynowe (A na rysunku) pośredniczą w poziomych oddziaływaniach, przenosząc informację między sąsiadującymi komórkami dwubiegunowymi i komórkami zwojowymi;
oraz komórki poziome (H na rysunku), które stanowią połączenia poziome w zewnętrznej siatkówce.
Zwróćmy teraz uwagę na drogę światła na rysunku 2. Światło musi przechodzić najpierw przez wszystkie pomocnicze komórki, zanim dotrze do fotoreceptorów — co na pierwszy rzut oka nie wydaje się być rozsądne. Jeśli problem konstrukcyjny, jakie każde oko musi rozwiązać, polega na ukształtowaniu maksymalnie dokładnego obrazu świata, to degradowanie światła, zanim osiągnie ono fotoreceptory, wydaje się w oczywisty sposób kiepskim rozwiązaniem. „Równoważne jest to umieszczeniu cienkiej półprzepuszczalnej zasłony bezpośrednio przed błoną filmową w twoim aparacie: może to tylko zmniejszać jakość obrazu”.[9] W tym właśnie miejscu ujęcia ewolucjonistyczne zaprzestają dalszej analizy.
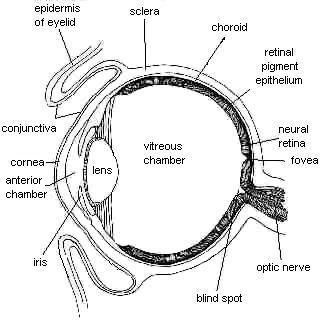
Rys. 1. Poziomy przekrój oka wyższych kręgowców. Istotną warstwą pomijaną przez ewolucjonistyczne dyskusje na temat optymalności siatkówki jest nabłonek barwnikowy siatkówki (RPE); patrz też Rys. 2. (Rysunek z: R. Adler and D. Farber, The Retina, Academic Press, New York 1986, s. 3.) |
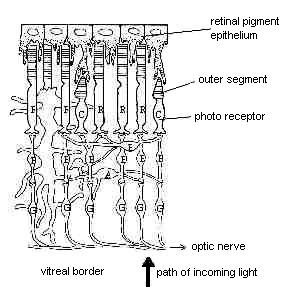
Rys. 2. Siatkówka dorosłego kręgowca. Na tym rysunku światło pada od dołu (czyli od ciała szklistego). Głównymi typami komórek są pręciki (R) i czopki (C), komórki poziome (H), komórki dwubiegunowe (B), komórki amakrynowe (A) i komórki zwojowe (G). Na górze rysunku znajduje się RPE, nabłonek barwnikowy siatkówki. Podobne do palców wyrostki RPE oddziałują z fotoreceptorami. (Rys. z: Adler and Farber, The Retina..., s. 5.) |
Istotna rola nabłonka barwnikowego siatkówki (RPE)
Należy jednak powiedzieć dużo więcej. Bezpośrednio przed siatkówką leży warstwa nabłonka utrzymująca fotoreceptory (patrz Rys. 2). Warstwa ta, zwana nabłonkiem barwnikowym siatkówki (odtąd RPE), jest istotna dla rozwoju i funkcjonowania siatkówki. I rzeczywiście, całe tomy poświęcono zrozumieniu roli RPE,[10] bowiem gdy źle on funkcjonuje, to źle funkcjonuje całe oko.
1. Regenerowanie barwników pochłaniających światło
Kiedy światło uderza w fotoreceptor, uruchamia łańcuch wydarzeń molekularnych, które ostatecznie kulminują się w ukształtowaniu obrazu w mózgu. Tutaj skupmy uwagę tylko na pierwszych etapach tego procesu.
Najpierw mamy do czynienia z wrażliwą na światło cząsteczką rodopsyny. Rodopsyna składa się z białka opsyny i innej cząsteczki, 11-cis-retinalu. Znajdująca się w odległych końcach fotoreceptorów — fragmencie najbliższym RPE, zwanym segmentem zewnętrznym — rodopsyna jest wbudowana w krążki błoniaste [utworzone przez wpuklone fałdy błony komórkowej — przyp. tłum.].[11] Kiedy światło uderza rodopsynę, energia zmienia kształt jej molekularnego składnika 11-cis-retinalu w całkowitą konformację trans. Proces ten nazywa się izomeryzacją. Ta konformacyjna zmiana w retinalu uruchamia złożoną kaskadę reakcji w kilku innych cząsteczkach, wywołując hiperpolaryzację (albo przesunięcie się ładunku elektrycznego) w krążku błoniastym zewnętrznego segmentu. Przekaźniki molekularne przenoszą wówczas ten sygnał elektryczny od synapsy u podstawy fotoreceptora do następnych neuronów, komórek poziomych i komórek dwubiegunowych — rozpoczynając w ten sposób proces, przy pomocy którego widzimy.
Proces ten zależy w istotny sposób od izomeryzacji 11-cis-retinalu. Każdy foton światła uderzający w fotoreceptor może izomeryzować retinal, a ponieważ wiele miliardów fotonów stale wpada do oka, retinal musi być regularnie zastępowany, by utrzymać ten cykl i ogólną funkcję fotoreceptora. Zadanie zastępowania przypada RPE. Komórki RPE zbierają zużyty retinal z fotoreceptorów i używają witaminy A do wyprodukowania świeżego retinalu, przenosząc go z powrotem do fotoreceptorów.[12]
2. Odzyskiwanie materiału fotoreceptywnego
Następną na liście funkcji, za które odpowiedzialny jest RPE, jest funkcja pokrewna: ponowne wykorzystywanie zużytych segmentów zewnętrznych. Krążki błoniaste segmentu zewnętrznego są bardzo aktywne i przez to muszą być stale zastępowane. Codziennie nowy krążek błoniasty segmentu zewnętrznego rośnie u podstawy tego segmentu (tam, gdzie krzyżuje się on z segmentem wewnętrznym, w rejonie komórki zawierającym jądro), zwiększając długość fotoreceptora. Gdy segment zewnętrzny oddala się od podstawy, jej odległy koniec — najstarszy krążek błoniasty — rozpada się na fragmenty. Fragmenty te są wychwytywane przez RPE, który fagocytuje ten materiał, wykorzystując ponownie wszystkie znajdujące się tam cząsteczki.[13] W ten sposób zużyte krążki błoniaste fotoreceptorów odrzucane są z optycznego toru światła i zastępowane przez nowy materiał. Ten stale zachodzący proces utrzymuje wysoką wrażliwość fotoreceptorów.[14]
3. Absorpcja nadmiernego światła
Oprócz tych aktywnych funkcji RPE pełni też ważną bierną rolę. Ponieważ jest on mocno zabarwiony, stanowi nieprzejrzysty ekran za fotoreceptorami. W ten sposób wchłania on światło nie odebrane przez fotoreceptory. Gdyby to się nie odbywało, światło to zmniejszyłoby rozdzielczość obrazów. Ta absorpcyjna własność RPE jest kluczowa w utrzymaniu wysokiej ostrości wzrokowej. To krótkie streszczenie nie wyczerpuje funkcji RPE. Zwróćmy na przykład uwagę, że RPE „jest niezbędne do normalnego rozwoju oka".[15] Funkcja ta nie jest bezpośrednio związana z widzeniem, ale z pewnością bez niej widzenie nie byłoby w ogóle możliwe. Krótko mówiąc:
Gdy rozważa się różne funkcje pełnione przez komórki RPE (...), nie ma cienia wątpliwości, że cały mechanizm metaboliczny RPE jest istotny dla normalnego funkcjonowania zewnętrznej warstwy siatkówki. Ze względu na naturę tych oddziaływań ważne jest, by RPE i fotoreceptory znajdowały się w bliskiej odległości.[16]
Istnieją ważne powody, by fotoreceptory kręgowców były skierowane tak, jak są.
Eksperyment myślowy
Ale nadal w każdej siatkówce znajduje się ślepa plamka. Oczywiście, ślepe plamki są przemieszczone poziomo względem siebie tak, że „gdy mamy oboje oczu otwartych, możemy widzieć wszystko, co się znajduje w polu widzenia”,[17] gdyż jedno oko widzi to, czego drugie nie widzi. Jednak możemy wyobrazić sobie sytuacje, w których to zawodzi:
Ślepe plamki naszych siatkówek rzadko kiedy powodują trudność, ale „rzadko” nie znaczy „nigdy”. Gdy na moment zamykam jedno oko chroniąc je przed owadem, jakieś ważne zdarzenie może się zogniskować na ślepej plamce drugiego oka.[18]
A więc w ramach eksperymentu myślowego zmieńmy ślepą plamkę. Zaczniemy od odwrócenia fotoreceptorów tak, aby ich przewody nie znajdowały się na drodze światła. Wyeliminowaliśmy w ten sposób ślepą plamkę, powodując nieco lepsze widzenie w jednym fragmencie oka. Teraz jednak naczynia krwionośne i RPE, potrzebne do utrzymania fotoreceptorów, muszą być umiejscowione po wewnętrznej stronie siatkówki, między nią a soczewką. A to ustawia sieć naczyń krwionośnych (zawierających wiele czerwonych ciałek krwi) oraz warstewkę nabłonka na drodze światła, znacznie zmniejszając informację wizualną docierającą do fotoreceptorów.
Ponadto ponieważ fotoreceptory stale zrzucają materiał ze swoich zewnętrznych segmentów, gromadzenie się tego nieprzezroczystego śmiecia na drodze światła w dużym stopniu zmniejszyłoby ilość światła docierającego do fotoreceptorów. Zaproponowana przez nas zmiana redukuje także jakość światła, gdyż ulega ono załamaniu na nieprzezroczystych kawałkach krążka błoniastego odrzuconego z zewnętrznego segmentu.
Możemy wyobrazić sobie, że po prostu umieszczamy RPE z tyłu siatkówki, ale wskutek tego powstanie problem, jak pozbyć się zużytych krążków błoniastych z zewnętrznych segmentów tak, aby fotoreceptory mogły się szybko zregenerować. Albo moglibyśmy otoczyć każdą komórkę fotoreceptora komórkami RPE, ale to wymagałoby zwiększenia miejsca między fotoreceptorami, zmniejszając w ten sposób rozdzielczość widzenia. Te projekty zmian wymusiłyby czasowe lub przestrzenne straty w widzeniu.
Czy są to więc ulepszenia? Z całą pewnością nie. W rzeczywistości nasz eksperyment myślowy spowodowałby, że oko kręgowców gwałtownie pogorszyłoby swoje parametry. Próbując wyeliminować ślepa plamkę, otrzymaliśmy wiele nowych i jeszcze gorszych funkcjonalnych problemów do rozwiązania. Nasza „naprawa” wydaje się dużo gorsza niż rzekoma wada, którą chcieliśmy usunąć.
Wniosek
Siatkówka kręgowców stanowi znakomity przykład funkcjonalnego — chociaż nie jest to intuicyjne — projektu. Projekt siatkówki jest odpowiedzialny za wysoką ostrość widzenia i wrażliwość na bodźce świetlne. Jest po prostu nieprawdą, że siatkówka jest w wyraźny sposób niedoskonała, ani nie jest łatwo wyobrazić sobie, jak mogłaby być zmodyfikowana bez znacznego pogorszenia funkcjonowania.
Dodatkowa literatura
H.S. Hamilton, The Retina of the Eye — An Evolutionary Road Block, Creation Research Society Quarterly, September 1985, Vol. 22, No. 2, s. 59-64.
H.S. Hamilton, Convergent Evolution — Do the Octopus ad Human Eyes Qualify?, Creation Research Society Quarterly, September 1987, vol. 24, No. 2, s. 82-85.
Tom Wagner, Darwin vs the Eye, Creation Ex Nihilo 1994, vol. 16, No. 4, s. 10-13.
Carl Wieland, Seeing back to front: are evolutionists right when they say our eyes are wired the wrong way?, Creation Ex Nihilo 1996, vol. 18, No. 2, s. 38-40.
An eye for creation. An Interview with eye-disease researcher Dr George Marshall, University of Glasgow, Scotland, Creation Ex Nihilo 1996, vol. 18, No. 4, s. 19-21.
Peter W.V. Gurney, Our 'inverted' retina — is it really 'bad design'?, Creation Ex Nihilo Technical Journal 1999, vol. 13, No. 1, s. 37-44.
Jerry Bergman, Is the Inverted Human Eye a Poor Design?, Perspectives on Science and Christian Faith, March 2000, vol. 52, No. 1, s. 18-30.
Peter Gurney, Dawkins' eye revisited, TJ 2001, vol. 15, No. 3, s. 92-99.
George Ayoub
Przypisy
* George Ayoub, Department of Biology, Westmont College, 955 La Paz Rd., Santa Barbara, CA 93108-1099.
[1] Autorem wprowadzenia jest zespół redakcyjny czasopisma Origins & Design
[2] William M. Thwaites, Design: Can We See the Hand of Evolution in the Things It Has Wrought?, w: Evolutionists Confront Creationists. Proceedings of the 63rd Annual Meeting of the Pacific Division, American Association for the Advancement of Science, San Francisco 1984, vol. 1, Part 3, s. 206-213; G.C. Williams, Natural Selection: Domains, Levels, and Challenges, Oxford University Press, Oxford 1992, s. 72-73.
[3] J. Diamond, Voyage of the Overloaded Ark, Discover, June 1985, s. 82-92; Richard Dawkins, The Blind Watchmaker, W.W. Norton, New York 1986 (wyd. polskie: Richard Dawkins, Ślepy zegarmistrz czyli, jak ewolucja dowodzi, że świat nie został zaplanowany, Biblioteka Myśli Współczesnej, PIW, Warszawa 1994, s. 155, 157); R.F. Miller, The physiology and morphology of the vertebrate retina, w: S.J. Ryan (ed.), Retina, 2nd ed. Volume 1: Basic Science & Inherited Retinal Disease, Mosby, St. Louis 1994, s. 58-71.
[4] Williams, Natural Selection..., s. 73.
[5] Por. tamże, s. 72.
[6] Por. tamże, s. 72.
[7] Miller, The physiology..., s. 30. Patrz także, Diamond, Voyage..., s. 91 oraz Williams, Natural Selection..., s. 74.
[8] K.D. Farnsworth and K.J. Niklas, Theories of optimization, form and function in branching architecture of plants, Functional Ecology 1995, vol. 9, s. 355 [355-363].
[9] T.H. Goldsmith, Optimization, Constraint, and History in the Evolution of Eyes, Quarterly Review of Biology 1990, vol. 65, s. 186 [181-322].
[10] Patrz na przykład R.H. Steinberg, Interactions between the retinal pigment epithelium and the neural retina, Documenta Ophthalmologia 1985, vol. 60, s. 327-346; K.M. Zinn and M.F. Marmor, The Retinal Pigment Epithelium, Harvard University Press, Cambridge, MA 1979.
[11] Istotna uwaga na temat terminologii, która może zmylić czytelnika: aktywny koniec komórki fotoreceptora, gdzie występują krążki błoniaste, nazywa się segmentem zewnętrznym. U kręgowców jednak ten segment faktycznie jest wewnętrzny, tj. znajduje się u spodu siatkówki i skierowany jest ku środkowi organizmu.
[12] Por. C.D.B. Bridges, Distribution of retinal isomerase in vertebrate eyes and its emergence during retinal development, Vision Research 1989, vol. 12, s. 1711-1717; A.T. Hewitt and R. Adler, The retinal pigment epithelium and interphotoreceptor matrix: Structure and specialized functions, w: Ryan (ed.), Retina..., s. 58-71.
[13] Por. D. Bok and R.W. Young, Phagocytic Properties of the Retinal Pigment Epithelium, w: Zinn and Marmor (eds.), The Retinal Pigment..., s. 148-174.
[14] Por. D. Bok, Retinal photoreceptor disc shedding and pigment epithelium phagocytosis, w: Ryan (ed.), Retina..., s. 58-71.
[15] S.M. Raymond and I.J. Jackson, The retinal pigment epithelium is required for the maintenance of the mouse neural retina, Current Biology 1995, vol. 5, s. 1286-1295.
[16] Hewitt and Adler, The retinal pigment..., s. 67.
[17] Williams, Natural Selection..., s. 73.
[18] Williams, Natural Selection..., s. 73.
Źródło: Na Początku... marzec-kwiecień 2002, nr 3-4 (153-154), s. 67-78. (George Ayoub, On the Design of the Vertebrate Retina, Origins & Design Winter 1996, vol. 17, No. 1, s. 19-22; z jęz. ang. za zgodą Autora tłum. Mieczysław Pajewski. Tłumacz składa podziękowanie za pomoc pani mgr Joannie Kaliszczyk)
