Homologia: gasnąca nadzieja neodarwinizmu
Wśród powodzi zapewnień o „fakcie ewolucji”, poczesne miejsce zajmuje homologia. Jest ona wymieniana jako jeden z najmocniejszych argumentów świadczących o prawdziwości koncepcji makroewolucji. Jednak, co dla wielu zaskakujące, pojęcie homologii wprowadzono przed Darwinem (rzecz jasna rozumiane morfologicznie, a nie filogenetycznie). Przeddarwinowska biologia wyjaśniała homologię przez odwołanie się do archetypów — spójnych wzorców, według których żywe organizmy zostały zaprojektowane. Karol Darwin, który przeformatował biologię w ujęcia naturalistyczne, zaproponował inne wyjaśnienie homologii — jako rezultat dziedziczenia z modyfikacjami od wspólnego przodka. Jednak by koncepcja ta była bardziej wiarygodna niż poprzednia, połączona być musi z empirycznie weryfikowalnym naturalistycznym mechanizmem odpowiadającym za powstanie homologii. Neodarwinizm proponuje dwa takie mechanizmy: program genetyczny i procesy embriogenezy. Jednak ich dokładniejsza analiza pokazuje, że żaden nie stanowi odpowiedniego dowodu. A wobec tego wyjaśnienie homologii odwołujące się do inteligentnego projektu wciąż pozostaje możliwą odpowiedzią.
Wśród powodzi zapewnień o „fakcie ewolucji”, poczesne miejsce zajmuje homologia. Jest ona wymieniana jako jeden z najmocniejszych argumentów świadczących o prawdziwości koncepcji makroewolucji. Jednak, co dla wielu zaskakujące, pojęcie homologii wprowadzono przed Darwinem (rzecz jasna rozumiane morfologicznie, a nie filogenetycznie). Przeddarwinowska biologia wyjaśniała homologię przez odwołanie się do archetypów — spójnych wzorców, według których żywe organizmy zostały zaprojektowane. Karol Darwin, który przeformatował biologię w ujęcia naturalistyczne, zaproponował inne wyjaśnienie homologii — jako rezultat dziedziczenia z modyfikacjami od wspólnego przodka. Jednak by koncepcja ta była bardziej wiarygodna niż poprzednia, połączona być musi z empirycznie weryfikowalnym naturalistycznym mechanizmem odpowiadającym za powstanie homologii. Neodarwinizm proponuje dwa takie mechanizmy: program genetyczny i procesy embriogenezy. Jednak ich dokładniejsza analiza pokazuje, że żaden nie stanowi odpowiedniego dowodu. A wobec tego wyjaśnienie homologii odwołujące się do inteligentnego projektu wciąż pozostaje możliwą odpowiedzią. Oczywiście nie dla ewolucjonistów, którzy wykluczają taką odpowiedź na podstawie przyjętych wcześniej filozoficznych założeń. Jednak zdroworozsądkowi sceptycy mogą sobie pozwolić spojrzeć na homologię chłodnym okiem, co też w tym artykule zrobimy.
Homologia morfologiczna i filogenetyczna
Od czasów Arystotelesa ludzie, którzy zajmowali się badaniami nad żywymi organizmami, stwierdzali niekiedy nadzwyczajne podobieństwa pomiędzy bardzo zróżnicowanymi stworzeniami. Nietoperze i motyle bardzo różnią się od siebie, ale co mają wspólnego, to skrzydła służące im do latania; nietoperze latają, a wieloryby pływają, ale układ kości w skrzydle nietoperza i w płetwie wieloryba są uderzająco podobne. Pierwszy rodzaj podobieństwa wymaga różnych struktur, które wykonują tę samą funkcję. W 1843 r. kreacjonistyczny biolog, Richard Owen, nazwał tego typu podobieństwa „analogią”. Dla kontrastu drugi rodzaj podobieństwa wymaga podobnych struktur, które wykonują zupełnie inne funkcje i Owen określił to zjawisko jako homologię. Owen i inni biologowie tego okresu wyjaśniali homologię istnieniem archetypów: biologiczne struktury są bardziej bądź mniej podobne, ponieważ zostały zaprojektowane według pewnych wzorców. [1]
W 1859 r. Karol Darwin zaproponował inne wyjaśnienie homologii. Zgodnie z tym wyjaśnieniem nietoperze i wieloryby posiadają podobną strukturę kości ponieważ odziedziczyły ją od wspólnego przodka, a nie dlatego, że były zaprojektowane zgodnie ze wspólnym projektem. Przez odrzucenie archetypów (co implikowało inteligentny projekt) i zastąpieniem ich hipotezą wspólnego pochodzenia opartą na naturalistycznym mechanizmie, Darwin miał nadzieję usunąć z terenu biologii koncepcje teleologiczne (czyli odwołujące się do celowości) i oprzeć ją na fundamencie naturalizmu — co też mu się z nawiązką udało.
Jednakże nie wszystkie podobne struktury są dziedziczone od wspólnego przodka (co wiedział już Darwin). Dla przykładu oko myszy jest strukturalnie podobne do oka ośmiornicy, pomimo, że ich hipotetyczny wspólny przodek nie posiadał oka tego typu. W 1870 r. Ray Lankester ukuł więc termin homoplazja do opisu podobnych struktur występujących w różnych gromadach, ale nie wskutek ich dziedziczenia od wspólnego przodka. Ewolucjoniści, wytresowani w dostosowywaniu swojej teorii do nowych faktów, wyjaśnili to zjawisko imając się ewolucji konwergentnej i paralelnej.
W tym rozróżnieniu kryło się jednak nowe niebezpieczeństwo. Jak stwierdził jeden z architektów neodarwinizmu, Ernst Mayr, po Darwinie „biologicznie najbardziej znaczącą definicją” homologii była następująca:
Cecha w jednym taksonie lub w większej ich ilości jest homologiczna, kiedy pochodzi ona od tej samej (lub odpowiadającej jej) cechy u ich wspólnego przodka. [2]
Innymi słowy to, co ewolucjoniści proponują jako wyjaśnienie homologii, staje się teraz jej definicją. Dla wielu biologów ta filogenetyczna definicja homologii zastąpiła jej strukturalną (morfologiczną) definicję. To z kolei niesie ze sobą poważne problemy natury logicznej, do czego jeszcze wrócimy.
Nowa koncepcja Darwina — wyjaśnienie homologii jao rezultatu dziedziczenia z modyfikacjami od wspólnego przodka — została w połowie XX w. ożeniona z nowo odkrytym mechanizmem genetycznego dziedziczenia, o którym Darwin nie wiedział nic. W ten sposób powstała synteza neodarwinowska, teoria mająca ambicję wyjaśniać znacznie więcej niż tylko istnienie homologii. Zasadniczym, klasycznym wyjaśnieniem homologii, jakie przedstawiał Darwin w XIII rozdziale O powstawaniu gatunków, jest dziedziczenie. Jak wiemy, każdy organizm posiada przynajmniej jednego rodzica. Tak więc — przykładowo — ludzie mają dwie duże kości w przedramieniu: łokciową i promieniową, ponieważ dziedziczą je od swoich rodziców, a ci z kolei odziedziczyli je od swoich rodziców itd. Cofając się w ten sposób w przeszłość możemy zakładać, że nasi odlegli człekokształtni przodkowie także posiadali takie dwie kości, choć najpewniej nieco inaczej ukształtowane. Ponieważ jednak taki wzorzec kości musiał pojawić się w jakimś określonym momencie ewolucyjnej historii, więc cofając się w czasie natkniemy się na hipotetycznego pierwszego przedstawiciela (wspólnego przodka), u którego po raz pierwszy wyewoluował taki wzorzec kości, od którego wywodzą się rozmaite grupy ssaków — wszystkie dziedzicząc ten układ.
Biologowie tworzący syntezę neodarwinowską dodali do tej opowieści Darwina nowy, fizyczny i przyczynowy wymir podstawy dziedziczenia. Mianowicie to, co jest przekazywane z pokolenia na pokolenie, to geny zakodowane w podwójnej helisie DNA. To właśnie geny sterują rozwojem zapłodnionego jaja, z którego poprzez proces embriogenezy powstaje dorosła postać danego organizmu. Ewolucja zaś występuje, ponieważ w genach zdarzają się mutacje, które mogą wywierać wpływ na rozwój i formowanie się organizmu, skutkiem czego w kolejnych generacjach powstają nieco zmienione fenotypy. Te fenotypy są następnie selekcjonowane przez dobór naturalny pod kątem przeżywalności i rozrodczości, czyli zdolności do wydawania potomstwa (zob. rys 1).
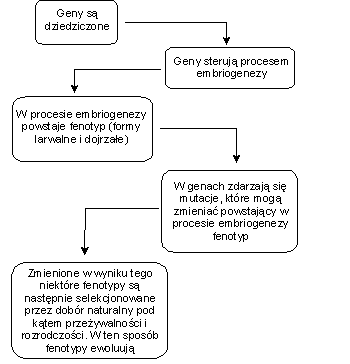
<!-- /* Font Definitions */ @font-face {font-family:"Cambria Math"; panose-1:2 4 5 3 5 4 6 3 2 4; mso-font-charset:0; mso-generic-font-family:roman; mso-font-pitch:variable; mso-font-signature:-1610611985 1107304683 0 0 159 0;} /* Style Definitions */ p.MsoNormal, li.MsoNormal, div.MsoNormal {mso-style-unhide:no; mso-style-qformat:yes; mso-style-parent:""; margin:0cm; margin-bottom:.0001pt; mso-pagination:widow-orphan; font-size:10.0pt; font-family:"Times New Roman","serif"; mso-fareast-font-family:"Times New Roman"; mso-ansi-language:PL;} p.MsoBodyText, li.MsoBodyText, div.MsoBodyText {mso-style-noshow:yes; mso-style-unhide:no; mso-style-link:"Body Text Char"; margin:0cm; margin-bottom:.0001pt; mso-pagination:widow-orphan; font-size:14.0pt; mso-bidi-font-size:10.0pt; font-family:"Times New Roman","serif"; mso-fareast-font-family:"Times New Roman"; mso-ansi-language:PL;} span.BodyTextChar {mso-style-name:"Body Text Char"; mso-style-noshow:yes; mso-style-unhide:no; mso-style-locked:yes; mso-style-link:"Body Text"; mso-ansi-font-size:14.0pt; mso-ansi-language:PL;} .MsoChpDefault {mso-style-type:export-only; mso-default-props:yes; font-size:10.0pt; mso-ansi-font-size:10.0pt; mso-bidi-font-size:10.0pt;} @page Section1 {size:612.0pt 792.0pt; margin:72.0pt 72.0pt 72.0pt 72.0pt; mso-header-margin:36.0pt; mso-footer-margin:36.0pt; mso-paper-source:0;} div.Section1 {page:Section1;} -->
Rys. 1. Schematyczne przedstawienie zasadniczych twierdzeń neodarwinizmu
Piękne i proste wyjaśnienie — w dodatku brzmi wiarygodnie. Obciążone jest jednak także poważnymi problemami. Szybki w ostatniej dekadzie rozwój badań nad genetycznymi mechanizmami embriogenezy, jak i ponowna analiza starszych, ale wciąż nierozwiązanych problemów, naruszyła fundament neodarwinistycznego wyjaśnienia homologii. Problemy tej koncepcji wyraził kilka lat temu David Cannatella z Departamentu Zoologii Uniwersytetu Teksańskiego pisząc:
Wake stwierdził, że homologia jest centralną koncepcją biologiczną. Jeśli jest to prawdą, to wielkiej grupie biologów porównawczych brakuje przewodniej koncepcji. Nie trzeba patrzeć daleko, by stwierdzić, że homologia (i zatem homoplazja) nie jest właściwie zrozumiana przez wielu biologów. [3]
Przyjrzyjmy się więc bliżej niektórym większym trudnościom neodarwinowskiego wyjaśnienia homologii, a zwłaszcza zagadkowymi przyczynowymi relacjami pomiędzy genami, embrionalnym rozwojem i formami fenotypowymi. Wbrew podręcznikowym zapewnieniom homologia nigdy nie była wyjaśniona przez neodarwinizm w sposób adekwatny. A skoro tak, należy ponownie poważnie przemyśleć zasadność wykluczenia z kręgu możliwych wyjaśnień inteligentnego projektu jako ewentualnej przyczyny istnienia homologii.
Ślepy Zegarmistrz pilnie potrzebny!
Gdy zapytacie znajomych ewolucyjnych biologów skąd wiedzą, że inteligentny projekt jest zbędny przy wyjaśnianiu homologii, usłyszycie mniej więcej coś takiego: „Znamy naturalistyczny mechanizm, który doskonale wyjaśnia ten fenomen”. W rzeczywistości jednak mechanizm ten nie został empirycznie potwierdzony, a inteligentny projekt odrzucono arbitralnie na podstawie filozoficznych restrykcji. Problem ten doskonale, choć nieświadomie i wbrew swym intencjom, naświetlił ewolucyjny biolog, Tim Berra w swojej książce z 1990 r. Evolution and the Myth of Creationism (Ewolucja i mit kreacjonizmu). Zgadzając się z nim:
Jeśli patrzysz na model samochodu Corvette z 1953 r. i porównujesz go z modelem dużo późniejszym, widoczne jest tylko ogólne konstrukcyjne podobieństwo, ale jeśli porównasz modele Corvette z roku 1953 i 1954 postawione obok siebie, a następnie modele z 1954 i 1955 r. pochodzenie z modyfikacjami będzie absolutnie oczywiste. To jest właśnie to, co paleontologowie robią ze skamieniałościami, dowody więc są tak mocne i obszerne, że nie mogą być odrzucone przez żadnego rozumnego człowieka. [4]
Jak wskazuje już sam tytuł książki, podstawowym celem Berry jest pokazanie, że żywe organizmy są rezultatem naturalistycznej ewolucji, a nie inteligentnego projektu. Jednak w swojej analogii Berra potwierdza to, co chciał skrytykować: Przyczyną strukturalnych podobieństw pomiędzy samochodami, jak i podobieństw pomiędzy starszymi a nowszymi modelami tego samego typu samochodu (o których Berra pisze „dziedziczenie z modyfikacjami”) jest istniejący wcześniej projekt. Analogia Berry znakomicie pokazuje, że nawet uderzające podobieństwa nie stanowią wystarczającego powodu do odrzucenia wyjaśnień bazujących na inteligentnym projekcie. Aby zademonstrować naturalistyczną ewolucję konieczne jest coś więcej. Należy empirycznie wykazać, że za mechanizmem, który odpowiada za powstanie nowych planów budowy ciała i nowych organizmów stoją przyczyny naturalistyczne i bezcelowe, czyli odmienne od tych, które odpowiadają za powstawanie kolejnych generacji samochodów.
Wielu naukowców nie chce jednak w ogóle dopuścić do alternatywy odwołującej się do inteligentnego projektu argumentując, że nauka ograniczona jest do badania li tylko naturalistycznych wyjaśnień. Chociaż to filozoficzne zastrzeżenie może wydawać się zasadne, to jednak nie może być ono kreowane jako dogmat, gdyż może stać się poważną przeszkodą w dociekaniu prawdziwych przyczyn wielu problemów. Ostatecznie nauka przez badania, formułowanie hipotez bazujących na empirycznych obserwacjach i eksperymentach ma prowadzić do bardziej adekwatnego i zgodnego z prawdą wyjaśniania naturalnych zjawisk. A to trochę coś innego niż twierdzenie, że nauka ograniczona jest tylko do naturalistycznych wyjaśnień tych zjawisk.
Zapewnienia, że coś jest obiektywnie prawdziwe — w naszym wypadku homologia wyjaśniana ewolucyjnie — bazować muszą na empirycznym materiale dowodowym, a nie tylko na wcześniejszych filozoficznych założeniach. Metodologiczne wykluczenie wyjaśnień odwołujących się do inteligentnego projektu wynika z ograniczeń tak zdefiniowanej nauki, a nie z empirycznego opisu obiektywnej rzeczywistości. Jeśli ewolucyjni biologowie chcą pokazać, że mechanizm ewolucji nie potrzebuje odwoływania się do inteligentnego projektu, nie mogą a priori wykluczać takiej możliwości, ale muszą poważnie potraktować to wyzwanie i precyzyjnie potwierdzić naturalistyczną alternatywę.
Alternatywa ta wyjaśniać musi to, co ewolucyjny biolog Leigh Van Valen określił „ciągłością informacyjną”. [5] Zgadzając się z nim, cechy homologiczne powstają podczas rozwoju każdego indywidualnego organizmu wskutek istnienia informacji, która jest z modyfikacjami dziedziczona od organizmu przodka. Tak więc pierwszym krokiem przybliżającym nas do zrozumienia mechanizmu ewolucji powinno być określenie natury informacji, która kontroluje embrionalny rozwój każdego organizmu.
Tańczący z genami: homologia i genetyka
Zasadniczą odpowiedzią na tak postawiony problem jest taka, że informacja ta zakodowana jest genach. W latach 30-tych XX w. synteza teorii Darwina z genetyką populacyjną pozwoliła określić zmiany (mutacje) w genach jako „motor ewolucji”, a kilka lat później odkrycie struktury i funkcji DNA rozszerzyło to wyjaśnienie na molekularny poziom. Zgodnie z neodarwinowską syntezą, informacja genetyczna zakodowana w DNA steruje embrionalnym rozwojem każdego organizmu. W procesie reprodukcji informacja ta przekazywana jest na następne pokolenia, ale mutacje w DNA (czyli zmiany w genetycznym programie sterującym rozwojem organizmu) czasem ją modyfikują (dziedziczenie z modyfikacjami), co może skutkować powstawaniem nieco zmienionych fenotypów (zob. rys. 1). Wskutek tego potomkowie wcześniejszego organizmu mogą posiadać struktury, które są podobne, ale nie identyczne (homologiczne). To ewolucyjne wyjaśnienie ma taką perswazyjną moc, że w latach 70-tych XX w. biolog molekularny Jacques Monod poczuł się uprawniony do obwieszczenia, że „mechanizm darwinistyczny jest co najmniej niekwestionowany” i że w konsekwencji „człowiek winien zrozumieć, że istnieje w wyniku zwykłego przypadku”. [6]
Jednak usiłowania skorelowania makroewolucji ze zmianami w sekwencjach genów nie zakończyły się jednoznacznym sukcesem. Dokładniejsze molekularne badania nie wykazały oczekiwanych związków pomiędzy mutacjami w genach i tym rodzajem fenotypowych zmian, który stałby się — jak to określił znany ewolucjonista — „surowcem ewolucji”. [7] Według Rudolfa Raffa i Thomasa Kaufmana:
Ewolucja napędzana mutacjami w DNA jest w znacznej mierze rozłączna z ewolucją na poziomie morfologicznym. (...) Najbardziej tego spektakularnym przykładem jest morfologiczna różnica pomiędzy ludźmi a szympansami pomimo tego, że ich DNA jest podobny w 99%. [8]
Część biologów w takiej sytuacji złożyła nadzieję w owym jednym procencie pozostałych genów, jakoby tu miała się kryć cała różnica pomiędzy człowiekiem a szympansem. Otóż wiadomo już od dawna, że nie wszystkie geny wykazują taką samą aktywność. Niektóre wydają się „uśpione”, bierne, inne wręcz przeciwnie, zaskakują swoją aktywnością. Co więcej — niektóre geny wydają się kluczowe dla prawidłowego rozwoju organizmu. Geny te sterują ekspresją całych grup innych genów wydając się w ten sposób nawigować wieloskalowe rozwojowe procesy. Geny te — nazywane homeotycznymi — determinują powstające w embrionalnym rozwoju poszczególne segmenty ciała organizmu i grają główną rolę w kontroli procesu rozwojowego. Doświadczenia laboratoryjne dowiodły, że nawet niewielkie mutacje w tych genach powodować mogą w rezultacie drastyczne zmiany w planie budowy ciała. Dla przykładu mutacja w homeotycznym genie muszki owocowej (Drosophila melanogaster) może spowodować transformację czułka tego owada w jego odnóże, lub wytworzyć dwie pary skrzydeł, podczas gdy normalnie powinna być tylko jedna. Inna mutacja w tego typu genach regulatorowych powoduje rozwijanie się oczu na odnóżach muszki.
Co istotne, geny niezwykle podobne do homeotycznych genów u muszek owocowych znaleziono u większości innych typów zwierząt włączając w to kręgowce. Wydają się one wspólne dla wszystkich eukariontów. Opierając się na tym, Eric Davidson i jego współpracownicy stwierdzili, że:
Nowe morfologiczne formy pojawiające się w ewolucji zwierząt są rezultatem zmian w genetycznym programie sterującym [embrionalnym] rozwojem. [9]
Zgodnie z tym spojrzeniem, cechy homologiczne są generowane przez homologiczne geny. Zakłada się, że jest nieprawdopodobne, by tak podobne homeotyczne geny wyewoluowały kilka razy niezależnie wskutek przypadkowych mutacji, tak więc ich rozpowszechnione podobieństwo oznacza, że poszczególne gromady zwierząt odziedziczyły je od wspólnego przodka. Podobne cechy generowane w różnych gromadach przez podobne geny stanowiłoby mocne potwierdzenie założenia o filogenetycznej — czyli ewolucyjnej — podstawie homologii. (Rzecz jasna, nie przeszkadza to ewolucjonistom ignorować rachunek prawdopodobieństwa i zakładać jednocześnie coś zupełnie przeciwnego — że ewolucja konwergentna i paralelna tworzyła niezależnie podobne struktury i geny wypełniające te same funkcje).
Jednak na tym klarownym obrazie — uniwersalności homeotycznych genów — zaczęły pojawiać się poważne pęknięcia. Okazało się bowiem, że owszem, muszki owocowe i myszy mają bardzo podobne homeotyczne geny, ale geny te sterują rozwojem zupełnie różnych narządów. Pomimo że myszy mają niezwykle podobne geny do tych, które — zmutowane — mogą powodować przekształcanie się czułki muszki owocowej w jej odnóże (antennapedia, muszki czółkonożne), u myszy nie rozwijają się ani czułki, ani odnóża, ale geny te sterują rozwojem jej móżdżka. I pomimo tego, że myszy i muszki mają bardzo podobne geny wpływające na rozwój oczu (muszki bezokie), wielofasetkowe oko muszki jest zupełnie odmienne od soczewkowego oka myszy. W obydwu przypadkach (muszek czółkonożnych i bezokich) homologiczne geny sterują rozwojem struktur, które nie są homologiczne ani wedle klasycznej morfologicznej definicji, ani wedle neodarwinowskiej filogenetycznej definicji. Podobny fenomen dotyczy genów ultrabithorax (Ubx), które u muszek owocowych uczestniczą w rozwoju skrzydeł, a ich odpowiednik u ludzi — gen homeobox (Hox) B7, zlokalizowany w chromosomie 17, bierze udział w procesach regulatorowych komórek, natomiast mutacje w tym genie mogą prowadzić do nieprawidłowego rozwoju nerek. Ewolucjoniści twierdzą, że te kompleksy genów musiały się pojawić u wspólnego przodka przed ewolucyjnym podziałem stawonogi/kręgowce. To jednak absolutnie nie wyjaśnia, gdzie tak bliźniaczo podobne pod względem struktury i położenia w chromosomach homeotyczne geny zawierają informację zdolną do rozwijania tak różnych i złożonych struktur (zob. rys. 2).
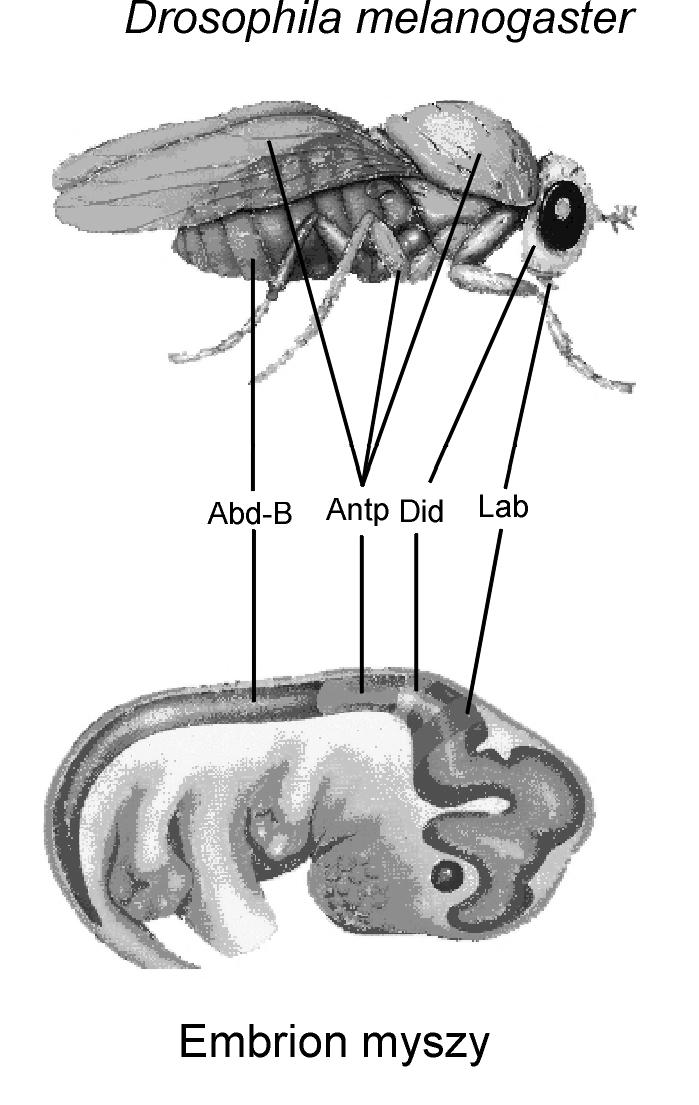
Rys. 2 Bliźniacze geny homeotyczne (Hom i Hox) występują u muszek owocowych i u myszy. Geny te — dziedziczone jakoby od wspólnego przodka, czyli homologiczne — sterują rozwojem niehomologicznych struktur.
Jeśli te same geny mogą „determinować” tak radykalnie różne struktury, znaczy to, że geny te tak naprawdę nie determinują struktur żadnych z nich. Są one konieczne do prawidłowego embrionalnego rozwoju, ale informacja w rzeczywistości odpowiedzialna za osiągnięcie właściwego rezultatu umiejscowiona musi być gdzie indziej. Spekuluje się, że geny te (homeotic selector genes) są swoistego rodzaju „włącznikami” kolejnych faz embriogenezy, czyli molekularnymi sygnałami, pod wpływem których organizm przechodzi do kolejnych etapów rozwojowej dynamiki. [10] Jednak „włączniki” czy też „sygnalizatory” to zupełnie coś innego niż „determinanty” zdolne do kształtowania fenotypowych struktur.
Co gorsza — nie tylko niehomologiczne struktury są rozwijane przez ponoć homologiczne geny, ale często zdarza się sytuacja odwrotna: niehomologiczne geny mogą rozwijać homologiczne struktury. Najbardziej znanymi tego przykładami, są — wspomniane wyżej — te wpływające na rozwój skrzydeł i oczu u muszek owocowych. Kiedy w embrionie muszki zmodyfikowano gen odpowiedzialny za rozwój skrzydeł, muszka rozwijała drugą parę skrzydeł dokładnie takich samych jak pierwsze, pomimo tego, że gen sterujący ich rozwojem był zmutowany. [11] Z kolei muszki, u których zmutowano gen sterujący rozwojem oczu, nie mogły ich rozwijać; ale jeśli muszki te były hodowane przez wiele kolejnych pokoleń, niektóre z ich potomków rozwijały prawidłowe oczy pomimo tego, że gen odpowiedzialny za ich rozwój był u nich cały czas zmutowany. Te anomalie doprowadziły embriologa Gavina de Beera do konkluzji, że: "Homologiczne struktury nie muszą być rozwijane przez identyczne geny" w związku z tym: "Dziedziczenie homologicznych struktur od wspólnego przodka (...) nie może być przypisane identycznym genom". [12]
Podstawowe założenie, że genetyczny program bezpośrednio kieruje embrionalnym rozwojem organizmów, zostało poważnie zakwestionowane przez wielu embriologów. [13] Sydney Brenner, który w 1970 r. pierwszy wprowadził pojęcie genetycznego programu, odrzucił później tę ideę, którą wcześniej rozwijał. Stwierdził on mianowicie, że informacja wymagana do formowania się połączeń nerwowych nawet u prostych owadów daleko wykracza poza informację zawartą w ich DNA. [14] Dekadę później embriolog, Brian Goodwin, stwierdził, że: "Geny determinują, jakie organizm wytwarza molekuły", jednak "Molekularna kompozycja organizmów zasadniczo nie determinuje ich form". [15]
W 1990 r. H.F. Nijhout w krytycznej analizie genetycznego programu konkludował, że: "Właściwym spojrzeniem na funkcje genów jest to, że zaopatrują one komórkę, i ostatecznie cały organizm, w chemiczny budulec. [16] Natomiast Wray i Abouheif podsumowali: "Homologiczne geny niekoniecznie kodują homologiczne struktury. [17]
Krótko mówiąc, wyjaśnienie homologii bazujące na genetyce okazało się nieadekwatne. Homologiczne geny mogą rozwijać niehomologiczne struktury, lub przeciwnie — niehomologiczne geny mogą rozwijać homologiczne struktury. Dostrzegając tę sytuację część biologów zasugerowała, że homologia jest rezultatem złożonych mechanizmów embriogenezy, których nie da się zredukować tylko do genetycznego programu.
Homologia i embriogeneza
Od kiedy okazało się, że homologia nie może zostać wyjaśniona przez samo tylko identyfikowanie informacji koniecznej do embrionalnego rozwoju zawartej w sekwencjach DNA, niektórzy biologowie spróbowali innej drogi: wyjaśnienie homologii przez przypisanie jej do podobnych rozwojowych procesów embriogenezy. Pomimo, że DNA determinuje sekwencje aminokwasów w białkach niezbędnych do rozwoju i funkcjonowania organizmu, na rozwój taki wpływa także szereg innych czynników, takich jak: lokalizacja składników cytoplazmy w jądrze komórkowym, fizyczne rozmiary embrionu i wiele innych. [18]
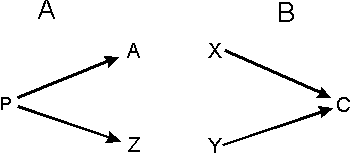
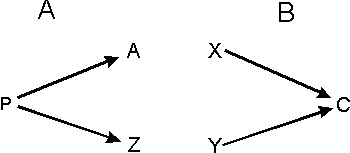
Rys. 3 Wspólne etapy embriogenezy (np. u różnych gatunków) mogą prowadzić do różnych końcowych form (A), lub przeciwnie, dwa różne rozwojowe szlaki mogą prowadzić do tej samej formy (B).
Jednak usiłowania skorelowania homologii z rozwojowymi procesami embriogenezy zakończyły się zupełnym niepowodzeniem. Po pierwsze, bardzo podobne rozwojowe procesy mogą prowadzić do zupełnie różnych cech. Jest dobrze znanym faktem, że na poziomie molekularnym praktycznie te same pozakomórkowe molekuły inicjujące poszczególne etapy embriogenezy mogą uczestniczyć w rozwoju niehomologicznych struktur u różnych zwierząt. [19] Na poziomie komórkowym wzorzec podziału komórek w rozwijającym się embrionie ptaków jest taki sam jak ten, w wyniku którego powstają niektóre gatunki żab. [20] I nawet na poziomie organizmów morfologicznie nierozróżnialne larwy mogły rozwijać się w kompletnie różne gatunki. [21] Podsumowując: niezwykle podobne rozwojowe procesy embriogenezy mogą prowadzić do zupełnie odmiennych rezultatów (zob. rys. 3).
Po drugie — i co bardziej zdumiewające — homologiczne cechy są często generowane wskutek działania zupełnie innych procesów rozwojowych. Nie ma wątpliwości, że jelito jest narządem homologicznym u wszystkich kręgowców pomimo tego, że formy jelit u różnych embrionów kręgowców są różne. Rdzeń kręgowy u niektórych strunowców formuje się z komórek będących prekursorem struny grzbietowej, podczas gdy u innych — nie. Najwyraźniej: "Struktury mogą zawdzięczać swoje pochodzenie różnym drogom bez tracenia swojej homologiczności. [22] Co więcej, jak stwierdził embriolog, Pere Albersch, jest „raczej normą niż wyjątkiem”, że "Homologiczne struktury formują się z wyraźnie odmiennych początkowych form. [23]
Powstawanie podobnych struktur formowanych w zupełnie różnych rozwojowych procesach jest także powszechne w późniejszych stadiach embrionalnego rozwoju. Wiele typów zwierząt przechodzi larwalne stadia w drodze do dojrzałości w fenomenie znanym jako niebezpośredni rozwój. Dla przykładu, większość żab rozpoczyna życie jako pływające kijanki, by później przejść metamorfozę w czworonożne zwierzę. Jest jednak także wiele gatunków żab, które omijają larwalne stadium kijanki i rozwijają się w dojrzałą formę bezpośrednio. I co zdumiewające, dojrzałe formy z tych, które rozwijają się bezpośrednio są w większości nie do odróżnienia od tych siostrzanych gatunków, które rozwijają się niebezpośrednio — tj. poprzez stadium kijanki.
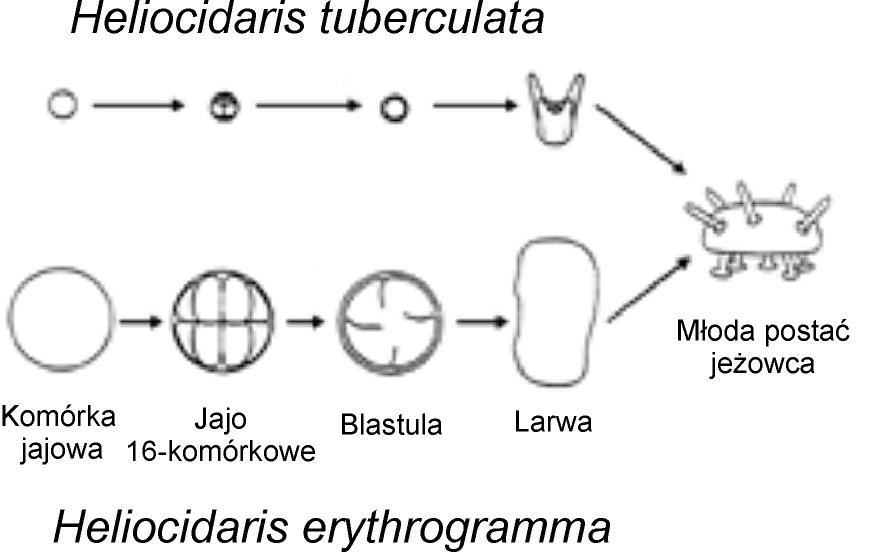
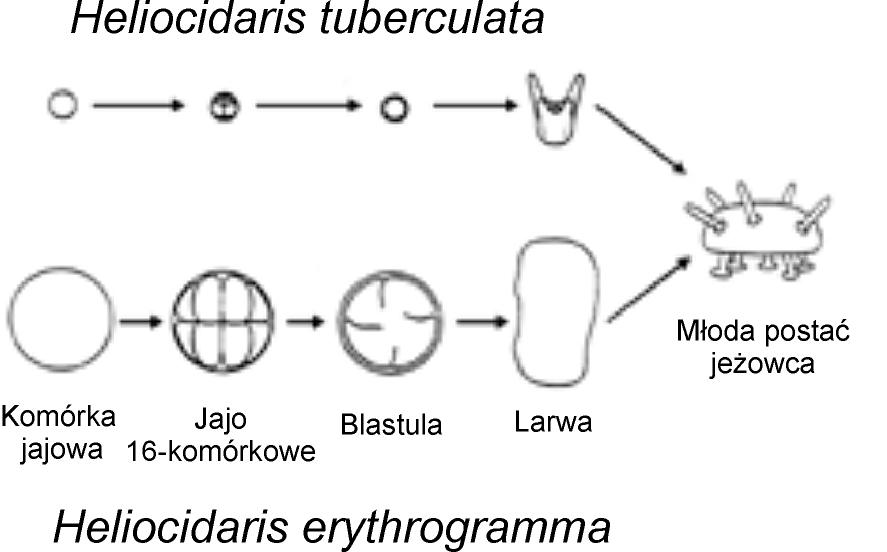
Rys. 4 Zupełnie różne ścieżki rozwojowe jeżowców prowadzą do prawie identycznych dojrzałych form.
Innymi słowy, mogą powstawać niezwykle podobne żaby pomimo tego, że ich drogi rozwojowe są radykalnie inne. Ten sam fenomen dotyczy jeżowców i żachw morskich (zob. rys. 4). [24]
Nawet klasyczny, podręcznikowy przykład kończyn kręgowców pokazuje, że homologia nie może zostać wyjaśniona przez odwołanie się do podobnych schematów rozwojowych w trakcie embriogenezy. Wzorzec kości w kończynach kręgowców powstaje w formie skondensowanej chrząstki, która później przekształca się w kość. Poszczególne etapy kondensacji tej chrząstki rozwijają się w procesie, który determinuje przyszły wzorzec kości w kończynach. Ale podobny wzorzec kości u różnych gatunków (tj. narządy homologiczne) wywodzi się z innych sekwencji kondensacji kostnej chrząstki. [25] Richard Hinchliffe podsumował to tak:
Embriologia nie wspiera porównawczej morfologii przez dostarczenie dowodów homologiczności kończyn w formie stałych wzorców tworzenia się zagęszczeń chrząstki wspólnych dla wszystkich tetrapodów. [26]
Tak więc embriologia nie zdołała rozwiązać problemu homologii. Podkreślić należy fakt, że embriony są w stanie wytwarzać niezwykle podobne struktury pomimo radykalnie różnych rozwojowych dróg, które do tych struktur wiodą. Naturalistyczne mechanizmy, póki co, nie są w stanie dostarczyć wyjaśnienia tego faktu, wyjaśnienia dostępnego do empirycznego badania. W 1958 r. Gavin de Beer stwierdził:
Relacje pomiędzy homologicznymi strukturami nie mogą być wyjaśnione przez odwołanie się do podobieństwa komórek w embrionie lub komórek jajowych znajdujących się poza miejscem, gdzie te struktury są ostatecznie formowane, lub rozwojowych mechanizmów, które budują te struktury. [27]
Następne pokolenia naukowców zdecydowanie potwierdziły poprawność obserwacji de Beera. Homologia, czy to definiowana morfologicznie, czy filogenetycznie, nie może być przypisana do homologicznej embriogenezy nie bardziej niż może być przypisana do homologicznych genów. Wniosek z tego, że naturalistyczne mechanizmy proponowane do wyjaśnienia homologii nie znajdują oparcia w faktach. Skoro tak, trzeba je przyjąć na wiarę — jest tak, ponieważ tak mówi teoria ewolucji. Spójrzmy, jak się to robi.
Kręcąc się w kółko
Wspominałem wyżej o innym problemie związanym z homologią, tym razem problemie leżącym w kwestii logiki jej zdefiniowania. Jak widzieliśmy, homologię można ująć rozmaicie, inaczej robiono to przed Darwinem, inaczej robi się to teraz. Ujęcia te można przedstawić w formie następujących sylogizmów:
Klasyczne (morfologiczne) ujęcie:
Przesłanka 1 (definicja): Narządy homologiczne to te, które posiadają podobne struktury.
Przesłanka 2 (empiryczna obserwacja): Skrzydła nietoperza i płetwy wieloryba posiadają podobne struktury.
Wniosek: Zatem skrzydła nietoperza i płetwy wieloryba są narządami homologicznymi.
Rozszerzenie klasycznej definicji przez Darwina:
Przesłanka 1 (wniosek z ujęcia klasycznego): Skrzydła nietoperza i płetwy wieloryba są narządami homologicznymi.
Przesłanka 2 (proponowane wyjaśnienie): Cechy są homologiczne, ponieważ są one dziedziczone od wspólnego przodka.
Wniosek: Zatem skrzydło nietoperza i płetwa wieloryba są to struktury odziedziczone od wspólnego przodka.
Ujęcie neodarwinowskie (filogenetyczne):
Przesłanka 1 (definicja): Cechy są homologiczne, jeśli są one odziedziczone od wspólnego przodka.
Przesłanka 2 (założenie): Skrzydło nietoperza i płetwa wieloryba są to struktury odziedziczone od wspólnego przodka.
Wniosek: A zatem skrzydło nietoperza i płetwa wieloryba są strukturami homologicznymi.
Jak widać, ujęcie neodarwinowskie jest klasyczną definicją w błędnym kole. Skąd wiemy, że dwie cechy są homologiczne? Ponieważ pochodzą one od wspólnego przodka. A skąd wiemy, że pochodzą one od wspólnego przodka? Ponieważ są one homologiczne. Zdumiewające jednak, że to wnioskowanie obciążone poważnym logicznym błędem powszechnie stosuje się do „dowodzenia prawdziwości” teorii ewolucji. I tak podręcznik Starra i Taggarta Biology: The Unity and Diversity of Life oświadcza, że:
Wzorzec makroewolucji — to jest zmienność od jednej formy wspólnego przodka — nazywany jest morfologicznym zróżnicowaniem (...) Homologia jest to podobieństwo jednej lub więcej części ciała u różnych organizmów wskutek dziedziczenia ich od wspólnego przodka. (...) Homologiczne struktury dostarczają bardzo mocnego świadectwa morfologicznego różnicowania się. [28]
Dokładnie tak samo stawiają sprawę Campbell, Reece i Mitchell:
Podobieństwo z charakterystycznym rezultatem wspólnego pochodzenia określane jest jako homologia, i takie anatomiczne znaki ewolucji nazywane są homologicznymi strukturami. [29]
Raven i Johnson w rozdziale „Dowody na makroewolucję są obszerne” również uczą studentów, jak definiować w błędnym kole: "Homologia: Wiele organizmów posiada organy, które są strukturalnie podobne do tych u ostatniego wspólnego przodka. To jest dowód ewolucyjnych pokrewieństw". Kilka stron dalej ten sam podręcznik jasno definiuje narządy homologiczne jako: "Struktury z różnym wyglądem i funkcjami wywodzące się od tych samych części ciała wspólnego przodka. [30]
Dowodzi to tylko, jak bardzo ewolucjonistyczne dogmaty potrafią zniekształcić ogląd rzeczywistości. Nie znaczy to, że strukturalne podobieństwa różnych narządów nie mogą być używane jako jeden z argumentów na wspólnotę pochodzenia, musi to być jednakże poparte innymi dowodami, np. paleontologicznym zapisem skamieniałych gromad stopniowo rozdzielających się od wspólnego przodka. Natomiast podobieństwo struktur samo w sobie nie uprawnia do wniosku o ewolucyjnej genezie tego podobieństwa.
Wnioski
Jak z powyższego widać, proponowane ewolucyjne wyjaśnienia zjawiska homologii okazały się dalece nieadekwatne. Próby skorelowania homologii z genami determinującymi rozwój organizmów i z procesami embriogenezy zakończyły się niepowodzeniem. Homologiczne geny mogą sterować rozwojem niehomologicznych struktur i przeciwnie — niehomologiczne geny mogą sterować rozwojem homologicznych struktur. Homologiczne procesy embriogenezy mogą wytwarzać niehomologiczne struktury i przeciwnie — niehomologiczne procesy embriogenezy mogą wytwarzać homologiczne struktury. Wszystkie kombinacje dozwolone.
Sytuację tę zaczynają dostrzegać niektórzy ewolucjoniści. Oto w artykule National Center for Science Education, którą John Rennie w swoim antykreacjonistycznym artykule w Świecie Nauki zachwalał jako „jedyną amerykańską organizację specjalizującą się w obronie nauczania teorii ewolucji przed atakami kreacjonistów” [31] stwierdzono:
Homologia nie jest „dowodem” na wspólnotę pochodzenia: wspólne pochodzenie jest wnioskiem bazującym na wielu innych źródłach informacji, wzmocnionym przez wzorzec podobieństw i różnic anatomicznych struktur. [32]
Czas najwyższy, by prawdę, że homologia nie stanowi dowodu na wspólne pochodzenie, wzięli sobie do serca także inni ewolucjoniści.
W 1902 r. William Paley napisał, że gdy ktoś przechodzący przez wrzosowisko ujrzy tam kamień, może racjonalnie wyjaśnić jego obecność naturalistycznymi przyczynami. Jednak gdy zobaczy zegarek i stwierdzi, że „jego różne części są dopasowane i celowo złożone”, jedyną sensowną konkluzją przechodnia będzie ta, że zegarek ten został zaprojektowany. Paley argumentował per analogiam, że racjonalny jest wniosek, że żywe organizmy także zostały zaprojektowane. [33]
W 1859 r. Karol Darwin przedstawił swoją koncepcję, z której wynikało, że wszystkie żywe organizmy są bardziej kamieniem z wrzosowiska Paleya niż zegarkiem z tegoż wrzosowiska i ogłosił, że wszystko, co Paley przypisał projektowi, może być wyjaśnione procesami naturalistycznymi — przez dziedziczenie z modyfikacjami. Jak jednak pokazała analogia Berry z samochodami, samo „dziedziczenie z modyfikacjami” nie wystarcza do wykluczenia projektu. Konieczne jest jeszcze wykazanie, że mechanizm dziedziczenia z modyfikacjami jest naturalistyczny — przeciwnie niż w analogii Berry, gdzie mechanizm ten opiera się na inteligentnym projekcie. Darwin sądził, że zaproponował taki mechanizm swoją teorią doboru naturalnego, ale — jak to widzieliśmy powyżej — nie udało mu się tego osiągnąć.
Różne organizmy posiadają homologiczne cechy. Homologia w niektórych przypadkach może, lub nie, istnieć z powodu dziedziczenia od wspólnego przodka, ale zdecydowanie nie występuje z powodu podobieństwa genów lub podobieństwa embrionalnych mechanizmów rozwojowych. W 1971 r. Gavin de Beer napisał:
Jaki działa tu mechanizm, który może wytwarzać homologiczne organy, te same „wzorce”, pomimo, że ich rozwój nie jest kontrolowany przez te same geny? Zadałem to pytanie w 1938 r. i od tej pory nie otrzymałem odpowiedzi. [34]
32 lata później odpowiedzi wciąż nie znamy. Skoro jednak nie znamy naturalistycznego mechanizmu mogącego wyjaśnić zjawisko homologii, neodarwiniści nie mają podstaw do twierdzenia, jakoby naukowo dowiedli, że żywe organizmy nie zostały zaprojektowane. Wciąż więc pozostaje możliwość — jak widzieliśmy to w doskonałej analogii Berry — że homologia jest rezultatem istniejącego wcześniej projektu. Nie potrafiąc przedstawić empirycznie sprawdzalnego mechanizmu większości biologów pozostaje tylko jedno wyjście: wykluczyć rozważania o projekcie na podstawie przyjętych a priori filozoficzno-metodologicznych założeń.
Twierdzą oni, że nauka jest ograniczona naturalistycznymi granicami wobec czego wyjaśnienia odwołujące się do inteligentnego projektu są poza jej zasięgiem. Homologia musi więc być wyjaśniana naturalistycznie nie bacząc na to, że w ten sposób wyklucza się rozważenie możliwego aspektu rzeczywistości. Kiedy skonfrontuje się ich z faktami, że — jak dotąd — takiego wyjaśnienia nie udało się zbudować, potwierdzają oni swoje metodologiczne oddanie składając wyznanie wiary, że pewnie taki naturalistyczny mechanizm zostanie któregoś dnia odkryty.
Być może będzie. Ale co, jeśli żywe organizmy rzeczywiście zostały zaprojektowane? Ktoś, kto znajdzie na łące zegarek i zacznie zastanawiać się nad jego pochodzeniem, popełni poważny błąd odrzucając a priori możliwość, że powstał on w wyniku inteligentnego projektu. Osoba taka może zmarnować całe swoje życie próbując tworzyć fałszywe wyjaśnienia. Jeśli nauka ma szukać prawdziwych odpowiedzi, to jest to przedziwny sposób uprawiania nauki.
To tak jak w starym dowcipie, kiedy wieczorny spacerowicz spotyka pijaka szukającego coś pod jedyną palącą się lampą na ciemnej ulicy.
— Co pan robi? — pyta spacerowicz.
— Szukam mojego zegarka — odpowiada pijak.
— Jest pan pewien, że zgubił go pan tutaj?
— Nie, zgubiłem go po drugiej stronie ulicy, ale tu jest przynajmniej jasno.
Niestety, postawa większości biologów przypomina postawę tego szukającego zguby pijaka. Przyjmowanie bazujących na filozofii naturalistycznych założeń ograniczających poszukiwania możliwych przyczyn homologii nie jest najlepszą postawą w badaniach nad życiem.
Michał Ostrowski
Przypisy
[1] Alec L. Panchen, "Richard Owen and the Concept of Homology", w: Brian K.Hall (ed.), Homology: The Hierarchical Basis of Comparative Biology, Academic Press, San Diego 1994, s. 21-62. Por. także Peter J. Bowler, Evolution: The History of an Idea, revised edition, University of California Press, Berkeley 1989.
[2] Ernst Mayr, The Growth of Biological Thought, Belknap Press, Cambridge MA 1982, cyt. za: Wells and Nelson, Homology..., s. 13
[3] David Cannatella, Review of Homology and Homoplasy, Systematic Biology 1997, vol. 46, s. 369 [366-369].
[4] Tim M. Berra, Evolution and the Myth of Creationism. A Basic Guide tothe Facts in the Evolution Debate, Stanford University Press, Stanford CA 1990, s. 117.
[5] Leigh M. Van Valen, Homology and Causes, Journal of Morphology 1982, vol. 173, s. 305-312.
[6] Cyt. za: Horace Freeland Judson, The Eighth Day of Creation, Simon & Schuster, New York 1980, s. 217.
[7] R.C. Lewontin, The Genetic Basis of Evolutionary Change, Columbia University Press, New York 1974, s. 160.
[8] Rudolf A. Raff and Thomas C. Kaufman, Embryos, Genes, and Evolution, Macmillan, New York 1983, s. 67, 78
[9] E.H. Davidson, K.J. Peterson and R.A. Cameron, Origin of Bilaterian Body Plans: Evolution of Developmental Regulatory Mechanisms, Science 1995, vol. 270, s. 1319 [1319-1325].
[10] Por. B. Alberts, D. Bray, J. Lewis, M. Raff, K. Roberts, J.D. Watson, Molecular Biology of the Cell, 3rd Edition Garland Publishing Inc., New York 1994, s. 1093.
[11] Zob. Brian K. Hall, Evolutionary Developmental Biology, Chapman & Hall, London 1992.
[12] Gavin de Beer, Homology: An Unsolved Problem, Oxford University Press, London 1971, s. 15-16.
[13] Por. Jonathan Wells, The History and Limits of Genetic Engineering, International Journal on the Unity of the Sciences 1992, vol. 5, s. 137-150.
[14] Sydney Brenner, The Genetics of Behaviour, British Medical Bulletin 1973, vol. 29, s. 269-271.
[15] Brian C. Goodwin, What Are the Causes of Morphogenesis?, Bioessays 1985, vol. 3, s. 32, 36.
[16] H.F. Nijhout, Metaphors and the Role of Genes in Development, Bioessays 1990, vol. 12, s. 441-446.
[17] G.A. Wray, and E. Abouheif, When is Homology not Homology? Current Opinion in Genetics & Development 1998, vol. 8, s. 675 [675-680].
[18] Por. Wells, The History and Limits..., s. 137-150.
[19] Zob. Scott F. Gilbert, Developmental Biology, 4th ed. Sinauer Associates, Sunderland MA, 1994.
[20] Por. Richard P. Elinson, Change in Developmental Patterns: Embryos of Amphibians with Large Eggs, w: Rudolf A. Raff and Elizabeth C. Raff, (eds.), Development as an Evolutionary Process, vol. 8, Alan R. Liss, New York 1987, s. 1-21.
[21] Zob. Gavin de Beer, Embryos and Ancestors, 3rd ed. Clarendon Press, Oxford 1958.
[22] de Beer, Embryos and Ancestors..., s 151
[23] Pere Alberch, Problems with the Interpretation of Developmental Sequences, Systematic Zoology 1985, vol. 34, s. 46-58.
[24] Zob. Rudolf A. Raff, The Shape of Life: Genes, Development, and the Evolution of Animal Form, The University of Chicago Press, Chicago 1996.
[25] Por. Neil H. Shubin, The Implications of „The Bauplan” for Developmentand Evolution of the Tetrapod Limb, w: J.R. Hinchliffe, J.M. Hurle, and D. Summerbell, (eds.), Developmental Patterning of the Vertebrate Limb, Plenum Press, New York 1991, s. 411-421.
[26] Richard Hinchliffe, Towards a Homology of Process: Evolutionary Implications of Experimental Studies on the Generation of Skeletal Pattern in Avian Limb Development, w: J. Maynard Smith and G. Vida (eds.), Organizational Constraints on the Dynamics of Evolution, Manchester University Press, Manchester 1990, s. 121 [119-131].
[27] de Beer, Embryos and Ancestors..., s. 152.
[28] Cecie Starr and Ralph Taggart, Biology: The Unity and Diversity of Life, 8th Edition, Wadsworth Publishing Company, Belmont, CA, 1998, s. 318-319.
[29] Neil A. Campbell, Jane B. Reece and Lawrence G. Mitchell, Biology, 5th Edition, The Benjamin/Cummings Publishing Company, Menlo Park, CA, 1999, s. 424.
[30] Peter H. Raven and George B. Johnson, Biology, 5th Edition, WCB/McGraw-Hill, Boston 1999, s. 412 i 416.
[31] John Rennie, 15 odpowiedzi na nonsensowne tezy kreacjonistów, Świat Nauki wrzesień 2002, nr 9 (133), s. 72 [66-72].
[32] Alan Gishlick, Responses to Jonathan Wells's Ten Questions to Ask YourBiology Teacher, 28 November 2001, http://www.ncseweb.org/resources/articles/7719_responses_to_jonathan_wells3_11_28_2001.asp; podkreślenie dodane.
[33] Por. William Paley, Natural Theology, Reprinted in 1972, St. Thomas Press, Houston, s. 2.
[34] de Beer, Homology: An Unsolved Problem..., s. 16.
Źródło: Na Początku..., styczeń-luty 2003, nr 1-2 (164-165), s. 22-48. Opracowano na podstawie artykułów: Jonathan Wells and Paul Nelson, Homology: A Concept in Crisis, Origins and Design Fall 1997, vol. 18, No. 2, s. 12-19; Jonathan Wells, Inherit the Spin, Dicovery Institute, 15 January 2002; Jonathan Wells, Critics Rave Over „Icons of Evolution”: A Response to Published Reviews, Discovery Institute, 12 June 2002.
